この章は書きかけです。
翻訳開始:2007/12/12
「 * 」この印はそのセクションの翻訳は完了していないことを示す。
Chapter 15: Neurotransmitters
シナプスにおける化学的伝達は4つのステップに分けられる。2つが前シナプスでもう2つは後シナプスでおこる。
ステップ1:伝達物質の合成
ステップ2:伝達物質の貯蔵と放出
ステップ3:後シナプス細胞膜における受容体との伝達物質の相互作用
ステップ4:伝達物質のシナプス間隙からの除去
前章ではステップ2と3について考えた。それは、伝達物質の放出とどのように後シナプス受容体で相互作用するかのステップであった。今度は、化学的シナプス伝達の最初と最後のステップをみることにしよう。伝達物質として用いられる分子の合成と、シナプス活動の後にシナプス間隙からそれを除去するステップだ。
シナプス伝達に関する生化学的過程を詳しく考える前に、”化学伝達物質”が何を意味するかを明らかにしておくことは重要である。1930年代の初期に、Otto Loewi がカエルの心臓の迷走神経末端からアセチルコリン(ACh)が放出されることを示し、Henry Dale が"コリン作動性"と"アドレナリン作動性"の伝達に関する仕事を報告た後に、化学伝達物質の概念は親しみのあるものになった。コリン作動性(cholinergic)やアドレナリン作動性(adrenergic)という用語はAChやノルアドレナリン(あるいはアドレナリン)を神経伝達物質として使うニューロンを示すために導入された。それ以来、他の物質も伝達物質として働くことが発見された。さらに、Bernard Latz が1950年代にした量子的放出(14章をみよ)に関する仕事により、伝達物質として働く物質は、大抵はシナプスの小胞の中に蓄えられ、exocytosis(開口分泌)によって放出されるものと見なされている。にもかかわらず、神経伝達物質として認められているいくつかの物質は、exocytosisと同様に、シナプス間隙に細胞質から直接放出される。このように、神経伝達物質の概念は、ニューロンの細胞生物学と受容体の薬理学に関する新しい情報を取り入れるために常に修正されねばならなかった。
訳者注:本文でnorepinephrineやepinephrineと表記されているものはそれぞれノルアドレナリンとアドレナリンと訳した。(アドレナリンもエピネフリンも同じものです)
最初の近似として、伝達物質を一つのニューロンによってシナプスで放出される物質で、後シナプス細胞に特定のやり方で影響を及ぼすものと定義できる。後シナプス細胞というのはニューロンかあるいは効果組織のことで、後者はたとえば筋細胞や分泌腺のようなものである。生物学におけるその他の多くの操作的な概念のように、伝達物質の概念は正確ではない。典型的には神経伝達物質はホルモンとは異なる。後シナプス細胞は伝達物質の放出される場所に近接していて、ホルモンは血中に放出され離れた標的に作用する。ただし、伝達物質は放出場所からいくらか離れた場所の標的に作用することはできる。 伝達物質はautocoidsとも異なっている。伝達物質が放出されたニューロンとは異なる標的に典型的には働くのに対し、autocoidは放出された細胞に作用する。そうではあるが、一部のシナプスでは、承認された伝達物質多くが後シナプス細胞の受容体だけでなく、伝達物質が放出された末端にある受容体、自己受容体(autoreceptor)も活動させる。Autoreceptorはたいてい進行中のシナプス伝達を調節する。例えば伝達物質のさらなる放出を制限数瑠事によって。神経伝達物質の重要な特徴は、その効果は過渡的であり、数ミリ秒から数分にわたって続くことである。にもかかわらず、神経伝達物質の活動は標的細胞に数時間から数日続く長期的な変化を与える。
包括的な定義に達するこれらの困難さにもかかわらず、限られた数の低分子量基質が一般的に神経伝達物質として受け入れられている。しかし、これらの基質でさえ、しばしば伝達物質機能を特定のシナプスで示すのが難しい。これらの困難さ故に、多くの神経生物学者は、以下の4つのクライテリアを満たさない限り、神経伝達物質とは認めない。
1.ニューロンで合成されること
2.前シナプス末端に存在し、十分な量が放出されることで後シナプスニューロンまたは効果組織(effector organに)に決められた活動を引きおこすこと
3.外から(薬として)適当な濃度で投与されたとき、内因性で放出された伝達物質の活動を厳密に模擬すること(例えば後シナプス細胞の同じイオンチャンネルやセカンドメッセンジャー経路を活性化する)
4.活動の場所(シナプス間隙)からそれを除去する特異的な機構が存在すること
神経系は、主として二種類の化学物質を伝達に使用している:低分子伝達物質と、神経活性ペプチドである。ペプチドとはアミノ酸がつながった短い鎖のこと。そちらの種類も大小の小胞(vesicle)の中に含まれている。神経ペプチドは大きな有芯小胞(large dense-core vesicle)に包まれており、分泌線や肥満細胞(mast cell)で見られるのと同様のエクソサイトーシス機構で中身が放出される。低分子伝達物質は小さな輝いた小胞(small lucent vesicle)に包まれていて、特定のCa2+チャンネルに密接に関連したアクティブゾーンにおけるエクソサイトーシスを通して中身を放出する(14章をみよ)。大きな有芯小胞は神経ペプチドと同様に低分子伝達物質も含むことが出来る。ほとんどのニューロンが両タイプの小胞をもっているが、比率が異なる。小さなシナプス小胞はアセチルコリン、クルタミン、GABA、グリシンを伝達物質として使うニューロンに特徴的である。一方で、大きな有芯小胞は、カテコールアミン作動性(catecholaminergic)ニューロンとセロトニン作動性(serotonergic)ニューロンで典型的である。副腎髄質(adrenal medulla)はしばしばエクソサイトーシス研究のモデルとして使われるが、副腎髄質は大きな有芯小胞と同様の分泌顆粒(secretory granule)のみを含んでいる。有芯小胞は、低分子伝達物質と神経ペプチドの両方を含むことが出来るので、共存伝達(cotransmission)において重要である。これについてはこの章の後の方で議論する。
9つの低分子量基質が一般的に神経伝達物質として受け入れられている。8つはアミンであり、このうち7つはアミノ酸かその誘導体である (Table 15-1)。9番目はATPかその代謝物である。アミン化学メッセンジャーは多くの生化学的性質を共有する。すべて、荷電した低分子であり、比較的短い生合成経路で形成される。また、中間代謝(intermediary metabolism)の主要な炭水化物基質から誘導される前駆体から合成される。中間代謝の他の経路のように、これらの神経伝達物質の合成は酵素によって触媒される。それはほとんど例外なく細胞基質である。ATPはミトコンドリアで生まれ、細胞中に豊富に存在している
Table 15-1 低分子伝達物質とその生合成で鍵となる酵素
| 伝達物質 | 酵素 | 活動 |
|---|---|---|
| アセチルコリン (Acetylcholine) | コリンアセチル化酵素 (Choline acetyltransferase) | 特異的 |
| 生合成アミン (Biogenic amines) | ||
|
ドーパミン (Dopamine) ノルアドレナリン (Norepinephrine) アドレナリン (Epinephrine) セロトニン (Serotonin) ヒスタミン (Histamine) |
チロシン水酸化酵素 (Tyrosine hydroxylase) チロシン水酸化酵素 (Tyrosine hydroxylase) と ドーパミンβ水酸化酵素 (dopamine β-hydroxylase) チロシン水酸化酵素 (Tyrosine hydroxylase) と ドーパミンβ水酸化酵素 (dopamine β-hydroxylase) トリプトファン水酸化酵素 (Tryptophan hydroxylase) ヒスチジン脱炭酸酵素 (Histidine decarboxylase) |
特異的 特異的 特異的 特異的 特異性不確定 |
| アミノ酸 (Amino acids) | ||
|
γ-アミノ酪酸 (GABA; γ-Aminobutyric acid) グリシン (Glycine) グルタミン酸 (Glutamate) |
グルタミン酸脱炭酸酵素 (Glutamic acid decarboxylase) 一般的代謝の酵素操作 (Enzymes operating in general metabolism) 一般的代謝の酵素操作 (Enzymes operating in general metabolism) |
おそらく特異的 特異的経路未確定 特異的経路未確定 |
他の生合成経路のように、アミン伝達物質の全体的な合成は典型的にひとつの酵素反応に制御されている。酵素の制御は、しばしばある種類のニューロンに特徴的であり、たいていは他の種類の成熟したニューロンでは起こらない。
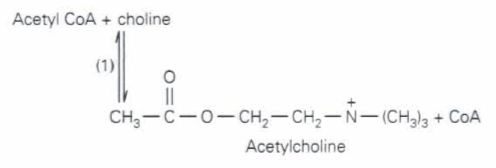
アセチルコリン(acetylcholine)は、受け入れられた低分子量アミン伝達物質であるが、アミノ酸でもその直接の誘導体でもない唯一のものである。ACh(アセチルコリン)の生合成経路は一つだけの酵素反応からなる。コリンアセチル化酵素(choline acetyltransferase)によってその反応は触媒される(下に示された反応のstep 1)。この転移酵素(transferase)はAChの生合成において特徴的なものである。神経組織はコリン(choline)を合成することはできない。コリンは食物から摂取され、血流を通してニューロンに運ばれる。共存基質(cosubstrate)であるアセチル補酵素A(acetyl CoA = acetyl coenzyme A)は多くの一般的代謝経路に関わっており、コリン作動性ニューロンに限られているわけではない。
アセチルコリンは脊髄の運動ニューロンで使われる伝達物質で、全ての脊髄動物神経筋接合部で放出される(11章)。自律神経系では、全ての神経節前ニューロンの伝達物質であり、副交感神経の神経節後ニューロンも同様である。脳全体の多くのシナプスで使われている。特に、AChを合成する細胞体は基底核(nucleus basalis)においておびただしく、大脳皮質へ広範な投射をしている。
ATPとその分解物(例:アデノシン adenosine)は一部のシナプスで伝達物質として働く。アデニン(adenine)とグアニン(guanine)とその分解物はプリン(あるいはピューリン purines)と呼ばれる;プリン作動性伝達の証拠は、特に精管(vas deferens)、膀胱(bladder)、心筋繊維への自律神経ニューロン;腸の平滑筋における神経叢(plexus);脳のニューロンの一部、において強い。プリン作動性伝達は特に痛みの生成において重要である(24章)。組織破壊によって放出されたATPは、ある種のイオンチャネル型のプリン受容体を通して、背根神経節の末梢C繊維軸索のむき出しの末端を興奮させる。背根神経節の中心軸索末端から放出されたATPは、脊髄後角にあるニューロンのもう一種類のイオンチャネル型プリン受容体を興奮させる。
ニューロンがどのように機能するかを研究する際の主要な仕事は、使われている化学メッセンジャーを同定することである。神経組織(nervous tissue)の組織断面における強力な組織化学(histochemical)技術で、低分子伝達物質と神経活性ペプチドの両方の探知することができる。
カテコールアミン(catecholamione)とセロトニン(serotonin)は、ホルムアルデヒド(formaldehyde)の蒸気と反応すると、蛍光誘導体(fluorescent derivative)を形成する。伝達物質の組織化学の初期の例では、スウェーデンの神経解剖学者のBengt FalckとNils Hillarpが、その反応を蛍光顕微鏡を適切に制御された条件下で用いることで伝達物質の場所を同定するために用いた。個々の小胞は光学顕微鏡の解像度では小さすぎて見えないので、組織蛍光(histofluorescence)は伝達物質の位置を神経細胞内で相対的で不正確にしか同定することが出来ない。(例えば細胞質か核か)正確な小胞の位置は、光学顕微鏡下での蛍光の分布と電子顕微鏡下での小胞の位置を比較することで調べられる。
組織化学解析は特別な条件下で、超構造(ultrastructural)レベルまで拡大することができる。過マンガン酸カリウム(potassium permaganate)かクロム酸塩(chromate)か銀塩(silver salt)の存在下での神経組織固定で、生合成アミンを含む小胞の電子密度が増大し、従ってアミン作動性ニューロンに特徴的な有芯(dense-core)小胞が多数現れる。
特定の伝達物質酵素やペプチド前駆体のための遺伝子を発現しているニューロンを同定することも可能だ。特定のmRNAを探知するためのたくさんの手法は、核酸のハイブリダイゼーション(hybridization)の現象に依存している。特にエレガントな手法の一つはin situ hybridization(インサイチューハイブリダイゼーション)である。核酸重合体の二本の単一鎖 は、もしその塩基配列が相補的(complementary)であればペアになる。in situ hybridizationを用いることで、非コーディングなDNA(noncoding DNA: 陰性(negative)あるいはアンチセンス(antisense)鎖、あるいはRNAに一致するもの)が、その内因性の(sense)mRNAとハイブリダイズするのに見合った条件下で組織断面に投与される。相補的な核酸鎖がmRNAと複合体を形成し、もしプローブ(核酸鎖)が放射線ラベルされていれば、オーロラジオグラフィー(autoradiography)によって、それを含むニューロンの場所が分かる。化学的なタグや免疫活性な塩基相同体を含むヌクレオチド(nucleotide)と一緒に合成されたオリゴヌクレオチド(oligonucleotide)が使われると、ハイブリッド(hybrid)は細胞化学的に局在化されうる。Figure15-3のように両方のラベルを同時に使うことが出来る。
Figure 15-3
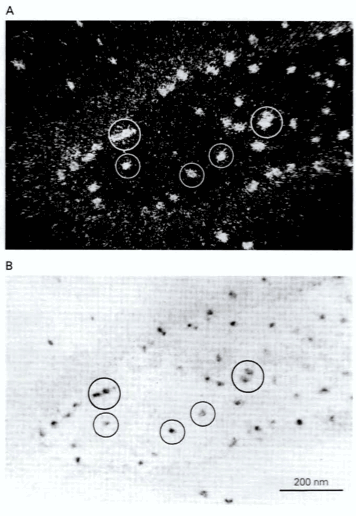
伝達物質は免疫細胞化学(immunocytochemistry)的にも位置を同定することができる。アミノ酸伝達物質、生合成アミン、神経ペプチドは、オートラジオグラフィーによってうまく位置を同定することが出来る。なぜなら、それらは第一級アミノ基(primary amino group)をもっており、これはニューロン内の位置に共有結合で固定をするのを可能するからだ;この官能基はアルデヒドによってタンパク質に架橋(cross-linked)する。アルデヒド(aldehyde)は顕微鏡観察で使われる一般的な固定液である。
製作:野澤真一(メールを送る)
http://melonsode.fem.jp/kandelyondel/